








公众号/ScienceAI(ID:Philosophyai)
编辑 | 萝卜皮
乳酸菌(LAB)是食品发酵中的重要微生物。在食品工业中,噬菌体(噬菌体或细菌病毒)可能会导致依赖 LAB 的过程中断,从而导致产品不一致和经济损失。LAB 噬菌体使用不同的粘附装置来感染宿主,但宿主结合机制的整体情况仍然不完整。
在这里,法国国家科学研究中心(CNRS)和艾克斯-马赛大学(Aix-Marseille University)的研究人员确定了感染葡萄酒细菌 Oenococcus oeni 的两种溶解性 siphophage OE33PA 和 Vinitor162 的粘附装置的结构和拓扑结构。这些噬菌体具有不同组成和形态的粘附装置,并且可能使用不同的感染机制。
研究人员使用 AlphaFold2 来获得粘附装置组件的 3D 模型。利用对 LAB 噬菌体宿主结合机器体系结构的先验知识,重建了 OE33PA 和 Vinitor162 粘附装置的拓扑结构。虽然 OE33PA 在其庞大的粘附装置的组装中表现出原始结构,但 Vinitor162 在其长而扩展的粘附装置中包含几个碳水化合物结合模块。
总体而言,这些结果突出了 AlphaFold2 预测蛋白质结构的能力,并说明了其在噬菌体结构和宿主结合机制研究中的巨大潜力。
该研究以「Structure and Topology Prediction of Phage Adhesion Devices Using AlphaFold2: The Case of Two Oenococcus oeni Phages」为题,于 2021 年 10 月 14 日发布在《Microorganisms》。

由于噬菌体长尾的灵活性,研究虹吸结构很困难。因此,通常使用「分而治之」的方法来解决这个问题,并确定噬菌体尾部成分的 3D 结构,包括多蛋白宿主粘附装置。特别是 X 射线晶体学和冷冻电镜揭示了这种组装的原子细节,从而为噬菌体-宿主相互作用提供了分子基础。在这种情况下,被证明对蛋白质结构预测高度可靠的 AlphaFold2 是分析多域蛋白质的完美工具,例如形成 siphophage 粘附装置的蛋白质。
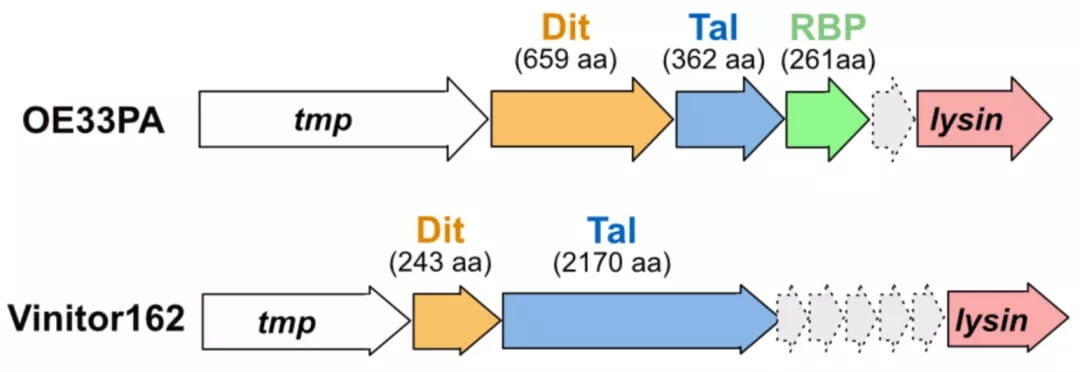
粘附装置编码基因组区域的示意图表示和分配。
宿主粘附装置是围绕共同支架构建的镶嵌组件,支架包括尾部远端的 Dit 六聚体环和附着在其上的 Tal 三聚体。这种 Dit-Tal 支架可作为适应专用 RBP 和辅助蛋白质的平台。模型乳球菌噬菌体 p2 包含 Dit、Tal 和 RBP,而其他噬菌体,包括乳球菌噬菌体 TP901-1 和 Tuc2009 以及葡萄球菌噬菌体 80α,组装了更多的成分。特别是,CBM 常见于粘附装置以及衣壳、颈部通道结构和尾管中,用于初步、可逆的糖类结合,涉及假定的宿主扫描。
这种直接反映在宿主结合机制中的组成多样性也存在于 O. oeni 感染噬菌体中:OE33PA 宿主粘附装置由 Dit、Tal 和 RBP 组成,而 Vinitor162 宿主粘附装置则只有两个核心组件。然而,Vinitor162 的 nsEM 成像揭示了一个高度灵活的 Tal 扩展,显示了几个被解释为 CBM 的灯泡,可能与宿主结合有关。
总体而言,OE33PA 粘附装置与众所周知的乳球菌噬菌体具有相似之处。值得注意的是,它似乎是一种「可激活」的粘附装置,类似于乳球菌噬菌体 p2 和 1358,并且可能使用 RBP 和 CBM 与宿主特异性结合。
此外,令人惊讶的是,数量相当有限的结构「砖」如何能够生产出多样化和特定于宿主的粘附装置。这在 OE33PA RBP 和演变的 Dit 中得到了清晰的解释。RBP 由在乳球菌噬菌体 1358 中发现的肩结构域和在史库纳病毒属(前 936 组)和 P335 组或李斯特菌噬菌体的乳球菌噬菌体中发现的头部结构域形成。因此,如在 p2 和 1358 粘附装置中观察到的那样,RBP 与 OE33PA Dit-Tal 核心的连接可能会接合 Dit「臂」。
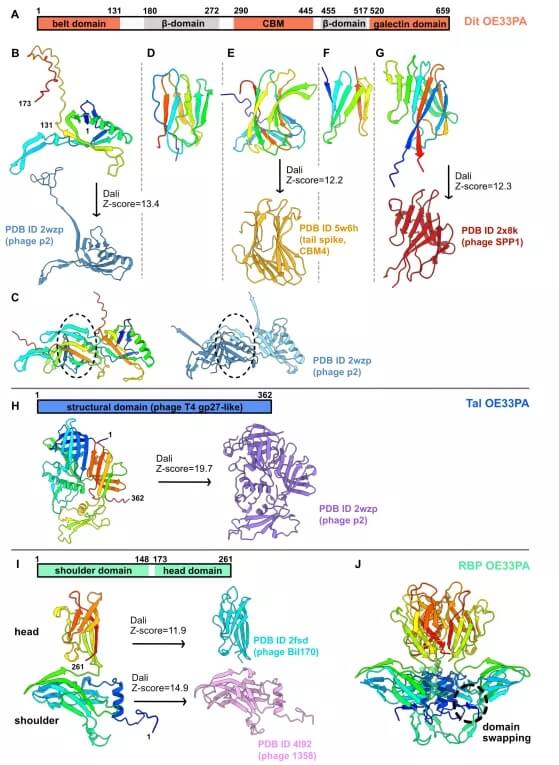
OE33PA Dit、Tal 和 RBP 的 AlphaFold2 结构预测。
关于进化的 Dit,它包含一个 CBM 插入,类似于 Skunavirus 属的许多其他噬菌体。值得注意的是,OE33PA Dit CBM 被预测为 CBM4,就像参与宿主细胞结合的噬菌体 Tuc2009(P335 组)的辅助蛋白 BppA 一样。然而,OE33PA Dit CBM 插入带和半乳糖凝集素结构域之间,两侧是 β-夹心结构域,而在进化出的 skunaviruses Dits 中鉴定的 CBM 插入 Dit「臂」内。与位于粘附装置外围的动态 Tuc2009 BppA 一样,OE33PA CBM 可能指向 Dit 环的外部。此外,该 CBM 通过一个长链接器连接到粘附装置的核心,这可能使其具有动态性并促进其与主机 CWPS 的交互。
Vinitor162 的 Dit 类似于噬菌体 p2 的 Dit,但没有「臂」延伸,而噬菌体 SPP1 的 Dit 也没有这种延伸。它不包含 CBM,如 OE33PA 和许多其他 skunaviruses 中所见。与 OE33PA 的 Tal 相比,Vinitor162 的 Tal 非常长,包含 2170 个残基。
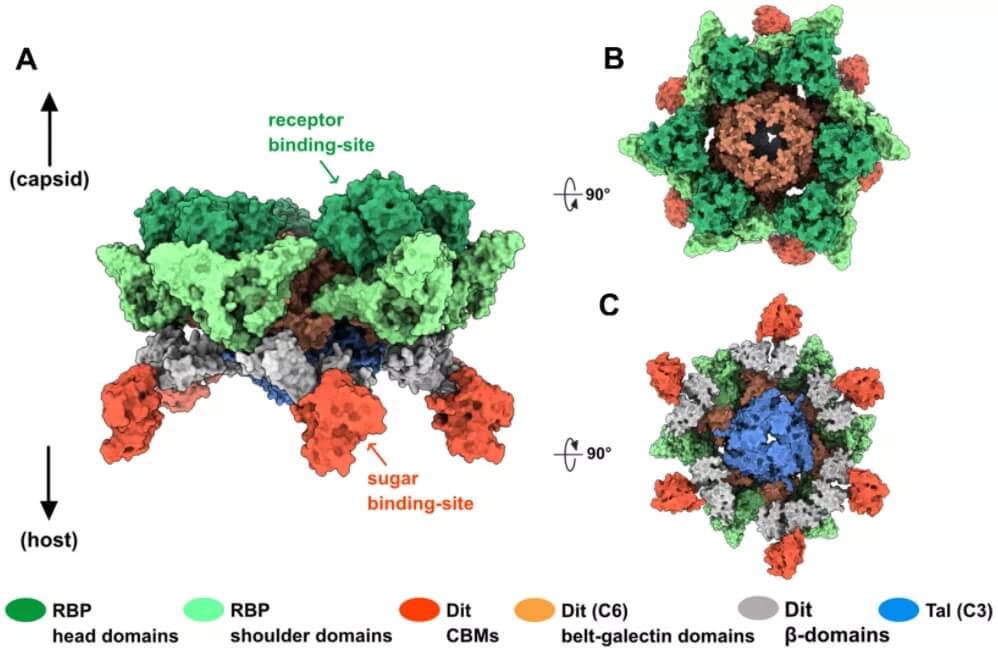
OE33PA 粘附装置的拓扑模型。
如此长的 Tals 并不罕见,之前在乳球菌噬菌体的 P335 组和嗜热链球菌噬菌体 pac 和 cos 中均有发现。用 HHpred 对这些长 Tals 的分析表明它们可能包含几个 CBM。AlphaFold2 预测了多达五个不同的模块,沿着 Tal 延伸和 Tal 尖端的 RBD,然后是伴侣域。五个模块中的四个被 Dali 明确识别为 CBM,而 CBM_2 不是,尽管它表现出 CBM 的经典 U 形。Tal 尖端类似于噬菌体 T5 L 形纤维的 C 端结构域,参与初步、可逆的噬菌体粘附到宿主。分子伴侣结构域确保纤维正确折叠,然后被 β-螺旋结构域切割。
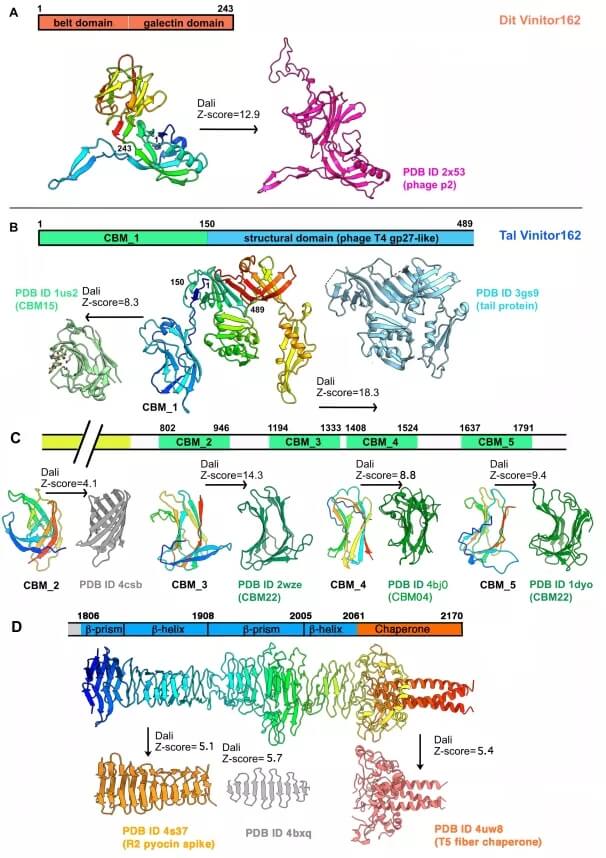
Vinitor162 Dit 和 Tal 的 AlphaFold2 结构预测。
Vinitor162 的 Tal 中的大量粘附模块非常显着,因为它占 15 个 CBM(每个 Tal 单体 5 个)和 3 个 RBD。然而,这 18 个粘附模块的总数与 skunavirus p2(6 个 RBP 三聚体)和 OE33PA 中的 RBP 数量相当。在 skunaviruses 中,这些 CBM 中的一些对其宿主的特异性是通过使用荧光标记的 CBM 的宿主细胞结合测定来确定的。在每种情况下,辅助 CBM 都表现出与真正的 RBP 相同的特异性。然而,OE33PA 的辅助 CBM 是否表现出与 RBP 相同的宿主特异性还有待确定。同样,不知道是否所有 Vinitor162 CBM 都具有功能,或者它们是否具有与 RBD 相同的宿主特异性。
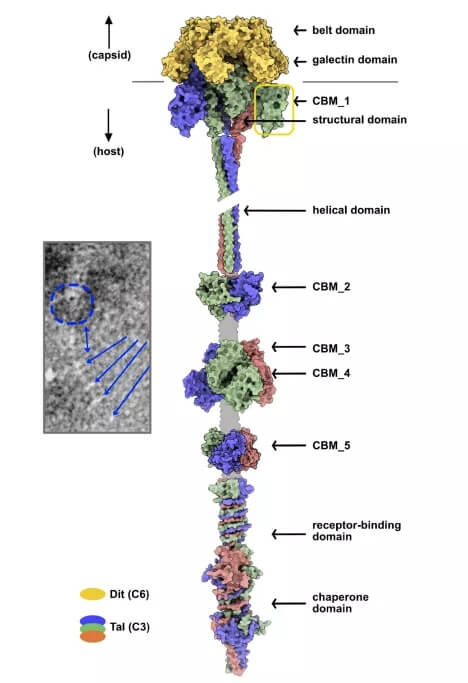
Vinitor162 粘附装置的拓扑模型。
在这里,AlphaFold2 以比 HHpred 更高的精度预测 CBM 域边界,这是这些域在大肠杆菌中成功重组表达和随后的荧光宿主细胞结合测定的先决条件。
论文链接:https://www.mdpi.com/2076-2607/9/10/2151/htm

 友情链接
友情链接